When you search 'flying bird step by step,' you are probably after one of two things: either you want to understand the actual biomechanics of how a bird moves from ground to air and back again, or you are chasing that deeper human itch to know what it would feel like to fly yourself. This article covers both. At its core, bird flight is a six-phase cycle: takeoff, climb, steady flapping, flap-glide and soaring, maneuvering, and landing. Each phase is driven by a precise sequence of muscular, skeletal, and aerodynamic events that evolution has refined over roughly 150 million years. Understanding that sequence, step by step, is one of the most satisfying journeys in all of biology.
Flying Bird Step by Step: How Birds Fly and How Humans Can

Marcus Chen
15 Jul 2026
What 'flying bird step by step' actually means (and who's asking)
The phrase sits at an interesting crossroads of intent. Some readers are students, naturalists, or curious adults who want the biomechanical play-by-play: which muscles fire first, how lift is generated, why a pigeon looks like it's struggling while a swift looks effortless. Others are drawn here by a more romantic impulse, the same one that shows up in songs, dances, and metaphors about freedom and escape. Both intentions are completely valid, and this site explores them seriously. The biological side is grounded in peer-reviewed research from journals like the Journal of Experimental Biology and synthesized in frameworks like Colin Pennycuick's landmark work on bird flight performance. The human side connects to everything from hang gliding to the cultural weight of flight as a symbol of liberation, something the related topic of what it feels like to fly like a bird explores in depth. See the related piece “How does it feel to fly like a bird” for a first-person, sensory exploration of human flight experiences and what they reveal about avian sensation.
One thing worth stating clearly at the outset: unaided human flight is biologically impossible. Our pectoral muscles make up roughly 1 percent of our body mass; a bird's pectoralis can account for 15 to 25 percent. Our bones are dense, our arms short, and we have no feathers to shape a wing surface. That does not mean you cannot experience genuine flight, but it does mean you need technology to do it. We will come back to those options after working through exactly how birds accomplish what we cannot.
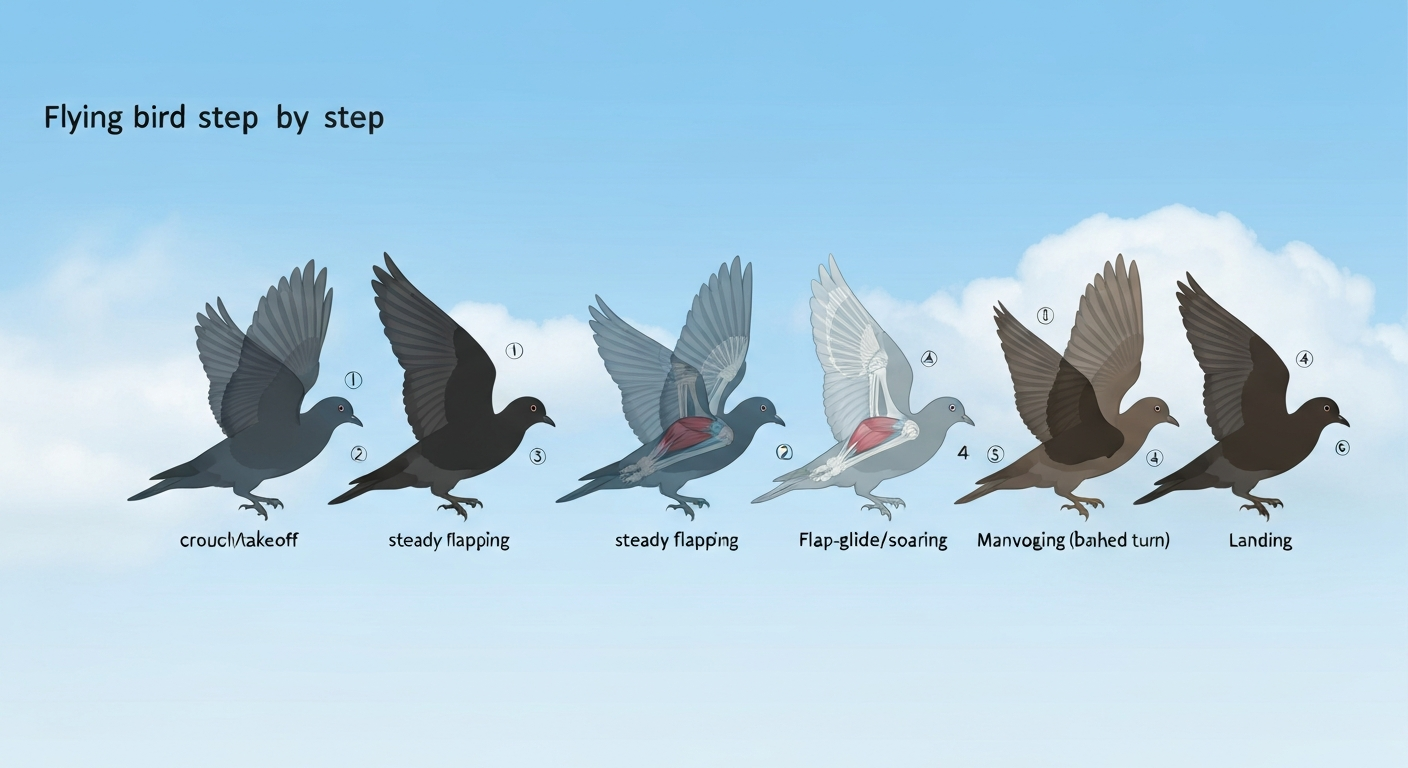
The six stages of bird flight at a glance
Bird flight researchers typically divide a complete flight sequence into six recognizable phases. The boundaries between them are not always sharp in real birds, but each phase has distinct kinematics, muscle activation patterns, and energy demands. Here is the sequence before we dig into each one.
| Stage | Phase Name | Key Action | Dominant Energy Mode |
|---|---|---|---|
| 1 | Takeoff | Jump + first downstroke, rapid wing extension | Very high (burst power) |
| 2 | Climb | Sustained thrust to gain altitude | High (aerobic + anaerobic) |
| 3 | Steady flapping | Continuous wingbeats at cruise speed | Moderate (sustained aerobic) |
| 4 | Flap-glide / soaring | Intermittent flapping or passive lift from thermals | Low to very low |
| 5 | Maneuvering | Asymmetric wing and tail adjustments | Variable (brief bursts) |
| 6 | Landing | Deceleration, drag deployment, controlled stall | Low mechanical output, high control demand |
Step 1: Takeoff
The sequence of events in those first fractions of a second
Takeoff is the most energetically expensive moment in a bird's entire flight. High-speed videography studies of pigeons show the sequence unfolds in under 200 milliseconds. The bird crouches slightly to load its leg muscles, then pushes hard against the substrate with both feet, generating a rapid upward and forward impulse. Simultaneously, the wings begin to extend from their folded resting position. The first downstroke starts before the feet leave the ground in many species, meaning the bird is already generating aerodynamic force while still in contact with the surface. In smaller birds like sparrows, the legs provide roughly 50 to 80 percent of the initial kinetic energy; in larger birds, wing power contributes proportionally more from the very first beat.
Wings, bones, and muscles doing the work
The downstroke is driven almost entirely by the pectoralis major, a massive chest muscle anchored to the keeled sternum (breastbone). This is the muscle that makes a chicken breast so large. In a pigeon, the pectoralis accounts for around 17 percent of body mass. The wing is extended at the shoulder, elbow, and wrist simultaneously, increasing the surface area just as it sweeps downward and forward. The upstroke relies on a cleverly engineered pulley system: the supracoracoideus muscle sits beneath the pectoralis but its tendon loops up through the foramen triosseum (a bony canal at the shoulder) to attach on top of the humerus. This arrangement means the supracoracoideus lifts the wing on the upstroke without requiring a muscle on the back of the body. The coracoid, furcula (wishbone), and scapula form a rigid tripod that braces against the enormous forces of each stroke.
The aerodynamics of getting airborne
At takeoff speeds, classical steady-state aerodynamics alone cannot fully explain how birds generate enough lift. Research published in the Journal of Experimental Biology has shown that leading-edge vortices (LEVs) play an important role during slow, high-angle-of-attack strokes. An LEV is a spinning column of air that forms along the leading edge of the wing when it moves at a steep angle through the air; it creates a low-pressure zone above the wing surface that augments lift well beyond what a flat-plate steady model would predict. This 'delayed stall' mechanism is the same one that keeps insects aloft and operates at the relatively low Reynolds numbers (roughly 10,000 to 100,000) characteristic of bird-scale wings moving at takeoff speeds. Modern fluid‑mechanics syntheses review unsteady aerodynamic mechanisms, steady lift, delayed stall/leading‑edge vortex (LEV), wake capture, induced drag, and vortex‑wake topology, and explain which models (quasi‑steady vs. vortex‑dominated CFD) apply across different size/Reynolds ranges (Vortices and forces in biological flight: insects, birds, and bats, Annual Review of Fluid Mechanics (review)) Vortices and forces in biological flight: insects, birds, and bats — Annual Review of Fluid Mechanics (review). The energy cost is steep: takeoff metabolic rates can reach 10 to 15 times a bird's resting metabolic rate for short bursts.
Step 2: Climb
Once airborne, the bird must gain altitude, and that requires generating more thrust than drag while maintaining enough lift to keep climbing. The wing beat frequency typically remains high during climb, close to or equal to takeoff frequency, and stroke amplitude may actually be slightly reduced as the bird settles into a rhythm. The tail fans slightly to increase surface area, contributing both lift and some pitch control. The supracoracoideus remains active to drive rapid upstrokes, because the bird needs to recycle the wing quickly for the next power stroke. From an energetics perspective, climb imposes a sustained demand on the aerobic oxidative muscle fibers (Type IIa and Type I) that are best suited for continuous contraction, while any anaerobic fast-twitch fibers recruited during the burst of takeoff begin to recover.
The ratio of thrust to lift required during a steep climb is substantially higher than during level flight. Pennycuick's flight performance models frame this as the bird operating near the top of its power curve, where mechanical power output is very high relative to the minimum power speed. A bird climbing at a steep angle is essentially paying an altitude 'tax' in metabolic currency. Field metabolic studies using doubly-labelled water have confirmed that migratory birds burn through their fat reserves fastest during the initial climb phases after departure, which is part of why many migrants time their departures to exploit favorable tailwinds from the very start.
Step 3: Steady Flapping
The stroke cycle broken down
Steady flapping cruise is the phase most people picture when they think of a bird in flight. Each wingbeat consists of a downstroke and an upstroke, and within each, the wing does not move as a rigid plank. During the downstroke, the primary feathers (the outermost, longest feathers attached to the 'hand' bones) twist to generate both lift and forward thrust simultaneously. The inner wing (attached to the 'arm' bones) acts more like a fixed aerofoil, generating predominantly lift. During the upstroke, the wing partially folds at the elbow and wrist in larger birds to reduce drag, while in smaller birds and at slower speeds, the upstroke can also be aerodynamically active, generating some lift through mechanisms including wake capture (where the wing intercepts the vortex wake left by the previous downstroke).
Muscle physiology and why it matters for endurance
The pectoralis in birds adapted for sustained flight is rich in mitochondria and myoglobin, giving it a distinctly dark red color (think duck or goose breast versus chicken breast). These aerobic fibers can sustain high-frequency contractions for hours by oxidizing fat. EMG studies by researchers including Bret Tobalske have shown that the pectoralis activates reliably at the beginning of each downstroke with a brief burst, while the supracoracoideus activates slightly before the end of the downstroke, pre-loading for the upstroke. This tight neuromuscular timing is one of the reasons birds can maintain precise wingbeat frequencies across a wide range of airspeeds.
The lift-thrust tradeoff and cruising speed
At any given speed, a bird is balancing four forces: lift against gravity, and thrust against drag. Faster flight increases lift (up to a point) but also dramatically increases induced and profile drag, so there is an optimal cruise speed where total power is minimized. Pennycuick's power curve models predict this 'minimum power speed' and a slightly faster 'maximum range speed,' and real birds have been shown via GPS and radar tracking to fly close to these theoretical optima during migration. A barn swallow cruising at around 9 to 11 meters per second and a migrating bar-tailed godwit maintaining roughly 15 meters per second are both close to their respective aerodynamic optima.
Step 4: Flap-Glide and Soaring
Intermittent flapping: working smarter, not harder
Many birds alternate bursts of flapping with brief glides or bound flight (wings folded entirely against the body). This is not laziness; it is a metabolic strategy. During the glide phase, the flight muscles can partially recover while the bird coasts on its kinetic energy. Research on swifts and swallows has shown that flap-gliding can reduce the overall cost of transport compared to continuous flapping, because the metabolic cost of a glide is very low while the aerodynamic cost (drag-induced speed loss) is modest over short intervals. Whether a species tends toward flap-gliding or bounding depends partly on wing loading (body mass divided by wing area). Swifts, with their long narrow wings and low wing loading, are excellent gliders. Woodpeckers, with higher wing loading and shorter wings, use deep undulating bounding flight instead.
Thermal soaring and why albatrosses almost never flap
Soaring takes two main forms. Thermal soaring, used by hawks, eagles, vultures, and storks, exploits columns of warm rising air (thermals) to gain altitude with almost no flapping. The bird circles within the thermal, trading slow circling drag for free altitude, then glides off in the desired direction, losing height slowly until it finds the next thermal. The key wing adaptation here is a high aspect ratio (long and narrow) wing or a broad wing with deeply slotted primary feathers (like a turkey vulture) that reduce induced drag during the tight-radius turns needed inside a thermal.
Dynamic soaring, the technique used by wandering albatrosses over the Southern Ocean, is even more elegant. The albatross exploits the wind gradient just above the ocean surface: it climbs into the wind (gaining energy from the wind shear), turns, and descends with the wind, skimming the wave faces, then repeats. GPS trajectory studies by Gottfried Sachs and colleagues have confirmed that albatrosses can travel thousands of kilometers on almost zero flapping power using this technique. Doubly-labelled water field studies have shown that a wandering albatross's flight costs are among the lowest of any bird relative to body mass, essentially comparable to sitting still. Their wingspan can reach 3.5 meters, and their wing loading and aspect ratio are specifically optimized for this wind-gradient exploitation.
Hovering: the most expensive mode of all
Hovering sits at the opposite end of the energy spectrum. Hummingbirds are the avian champions of stationary flight, beating their wings at 40 to 80 times per second and tracing a figure-eight stroke pattern in a nearly horizontal plane to generate lift on both the downstroke and a partially active upstroke. Kinematic work by Tobalske and colleagues using high-speed cameras and particle image velocimetry has shown that hummingbird wings generate wake vortex rings on each half-stroke, a pattern quite different from the single-loop wake of a bird in forward flapping flight. The metabolic cost of hovering in a hummingbird is roughly equivalent to a human running at full sprint, mass for mass.
Comparing flight modes across species
| Species | Primary Flight Mode | Wing Shape | Approximate Wing Loading (N/m²) | Notable Feature |
|---|---|---|---|---|
| Wandering albatross | Dynamic soaring | Long, narrow (high aspect ratio) | ~140 | Near-zero flapping cost over ocean |
| Ruby-throated hummingbird | Hovering + fast forward flight | Short, paddle-like | ~25 | Figure-eight stroke, 50+ beats/sec |
| Turkey vulture | Thermal soaring | Broad, slotted primaries | ~43 | Dihedral wing posture for stability |
| Peregrine falcon | High-speed dive (stoop) | Swept, pointed | ~75 | Exceeds 300 km/h in stoop |
| Common swift | Flap-glide cruising | Long, crescent-shaped | ~19 | Can remain airborne for 10+ months |
| Ostrich | Flightless (running) | Reduced, vestigial wings | N/A | No keeled sternum; largest living bird |
Flightless birds like ostriches, penguins, and kiwis represent a different evolutionary path entirely. Their sternal keels are reduced or absent, their pectoral muscles are proportionally small, and their wing bones are modified for other functions (penguins use theirs as flippers). Comparative anatomy studies have documented the correlation between keel reduction and loss of powered flight across multiple independent lineages. See the comparative analysis 'The relationship between sternum variation and mode of locomotion in birds, comparative/PMC article' (open‑access) for a synthesis of sternal keel reduction, wing‑loading changes, and other morphological correlates of flight loss across independent lineages blank" rel="noopener noreferrer">The relationship between sternum variation and mode of locomotion in birds — comparative/PMC article. The trade-off is almost always an advantage in some other ecological dimension: greater running speed, deeper diving ability, or access to a food resource that does not require flight to exploit. For another relevant comparison, see learn to fly like a bird.
Step 5: Maneuvering and Turning
Watching a peregrine falcon adjust its stoop trajectory by millimeters to intercept a starling, or a pine siskin weave through a dense spruce canopy at speed, you realize that straight-line flight is only part of the story. Maneuvering requires asymmetric adjustments: one wing extends slightly more than the other to generate differential lift, the tail rotates and fans asymmetrically to introduce a yawing or rolling moment, and individual primary feathers can be independently spread or tucked to fine-tune the wing's camber and surface area on each side.
The muscles responsible for these fine adjustments are not the massive pectoralis and supracoracoideus, but smaller muscles attached to individual feather follicles and to the wrist and finger bones of the wing. Electromyography studies have identified over a dozen discrete muscles involved in primary feather control alone. The tail is controlled by the levator and depressor caudae muscle groups, and its contribution to maneuvering is significant: birds with experimentally shortened tails in research studies show measurably reduced turning agility and longer pull-out distances from dives.
Aerodynamically, a banked turn works the same way in a bird as in an aircraft: the bird tilts its wing plane so that the total aerodynamic force vector has both an upward component (supporting weight) and an inward component (providing centripetal acceleration). Tighter turns require more bank angle, which means less vertical lift, which means the bird must also increase its speed or wingbeat frequency to avoid losing altitude. Agile species like accipiters (goshawks and sparrowhawks) have evolved relatively short, rounded wings that allow rapid changes in stroke geometry, at the cost of less efficient straight-line cruise.
Step 6: Landing
Slowing down without crashing
Landing is, in some ways, the most technically demanding phase. The bird must bleed off all of its forward momentum and kinetic energy in a controlled way, arriving at zero velocity precisely at the landing surface. The approach involves a progressive nose-up pitch attitude, which increases the wing's angle of attack and therefore drag. The tail fans fully to its maximum spread, acting as an aerodynamic brake. The alula (a small tuft of feathers on the bird's 'thumb') deploys forward to maintain airflow attachment over the main wing at high angles of attack, functioning exactly like the leading-edge slat on a commercial airliner. Without the alula, the wing would stall uncontrollably.
The kinematics of touchdown
Pigeon landing studies using high-speed cameras at the Journal of Experimental Biology level of analysis show that the bird executes a series of rapid, short, high-amplitude wingbeats in the final half-second before touchdown, with the wing swept increasingly forward and the body tilted steeply backward. The legs extend and the feet spread to maximize the contact area and absorb impact. The landing stroke generates a distinctive aerodynamic 'clap-and-fling' variant at very low speeds as the wingtips approach and the bound vortex is shed into the wake, producing the audible 'whoosh' you sometimes hear as a large bird settles onto a branch. Kinetic energy is dissipated both through aerodynamic braking and through elastic and muscular absorption in the legs. Unlike takeoff, landing does not require the bird to generate much net mechanical power outward; the energetic cost is more about precise neuromuscular control than brute force.
What birds sense during flight, and why it matters for landing
Birds have a remarkable suite of sensory tools that make precision landing possible. Their color vision extends into the ultraviolet range, giving them a richer perception of spatial detail than humans possess. Their vestibular system (the semicircular canals of the inner ear) is proportionally large and sensitive, providing rapid feedback on roll, pitch, and yaw that reaches the brain faster than human vestibular signals do. Mechanoreceptors (Herbst corpuscles) embedded in the bill and at the base of feather follicles detect pressure and airflow changes in real time, functioning essentially as a distributed airspeed and angle-of-attack sensor array. All of this feeds into a sensorimotor control loop that allows a bird to execute a 300-millisecond landing flare with a precision that no human-built autonomous vehicle has yet fully replicated.
Wing anatomy that makes all six steps possible
It is worth pausing to appreciate the anatomy that underlies every phase above, because the same structures appear again and again at each step. The avian wing skeleton is a modified forelimb: humerus, radius and ulna (fused at the wrist end), and a reduced set of fused hand and finger bones (the carpometacarpus and digits). The primary feathers attach to the hand bones and generate most thrust. The secondary feathers attach to the ulna and generate most lift. The scapula, coracoid, and furcula form the pectoral girdle that braces against stroke forces. The furcula (wishbone) acts as a spring, flexing outward during the downstroke and recoiling to assist the upstroke, storing and returning elastic energy somewhat like a bow.
- Humerus: upper arm bone; connects wing to shoulder; houses major muscle attachment points
- Radius and ulna: forearm bones; anchor secondary flight feathers along the trailing edge
- Carpometacarpus: fused wrist and hand bones; support primary feathers that drive thrust
- Alula ('thumb'): small leading-edge slot feathers; prevent stall at low speeds
- Pectoralis major: main downstroke engine; 15 to 25% of body mass in strong fliers
- Supracoracoideus: upstroke driver via pulley tendon through foramen triosseum
- Furcula (wishbone): elastic spring that stores and returns energy across each stroke cycle
- Tail feathers (rectrices): controlled by levator and depressor caudae; essential for steering and braking
Why humans cannot fly unaided, and what we can do about it
The numbers are unforgiving. A 70-kilogram human with arms spanning roughly 1.8 meters would need a wing area of about 7 square meters and a pectoral muscle mass of around 14 kilograms just to approach the minimum power requirements for sustained level flight, according to the basic scaling relationships in Pennycuick's models. Our actual pectorals weigh about 0.6 kilograms each. Our bones are solid and heavy. Our muscle fiber composition is not optimized for the high-frequency, high-force contractions that flight demands. No amount of training or wishing changes this fundamental biomechanical reality.
That said, the human desire to fly like a bird has produced some genuinely extraordinary technologies that let you experience the sensory and kinesthetic reality of flight in ways that would have seemed miraculous a century ago. If you want practical steps and technologies for experiencing avian-like flight, see our guide on how can I fly like a bird. Each of the following has a real on-ramp for a motivated adult.
| Activity | Closest Bird Analogy | Minimum Age / Requirements | How to Start |
|---|---|---|---|
| Paragliding | Thermal soaring (hawk, eagle) | 16+ in most countries; no prior experience needed | 2-day beginner course with certified school; tandem first flight available same day |
| Hang gliding | Dynamic gliding (albatross) | 14+ for tandem; fitness and coordination helpful | Tandem flight with instructor; solo certification takes 5-10 days of lessons |
| Wingsuit BASE jumping | Diving/gliding (swift, swift-like) | 18+; requires 200+ skydives as prerequisite | Skydiving license first; wingsuit progression course with experienced mentor |
| Skydiving (freefall) | High-speed dive/stoop (falcon stoop) | 16-18 depending on country | Accelerated Freefall (AFF) 8-jump course with dual instructors |
| Indoor skydiving (wind tunnel) | Hovering, body positioning | 3+ years old; no experience needed | Walk-in sessions at indoor skydiving centers; 1-2 minute flights typical |
Paragliding and hang gliding are the closest functional analogues to the soaring phase of bird flight: you are riding thermals and ridge lift using a wing that obeys the same aerodynamic principles that keep a red-tailed hawk aloft on a summer afternoon. Experienced paraglider pilots report the same kind of sensory feedback a bird uses, feeling subtle pressure changes through the wing and adjusting posture in response. The physics, if not the biology, is nearly identical.
Flight in culture: songs, dances, metaphors, and the fiction of shapeshifting
Flight as a metaphor for freedom, aspiration, and transcendence appears in virtually every human culture. The lyric bird flying high you know how i feel from 'Feeling Good' captures that emotional association. The phrase 'fly like a bird' echoes through everything from Nina Simone's 'Feeling Good' (those soaring lyrics about a new dawn and a bird flying high) to gospel traditions where flight represents liberation from earthly suffering. For a closer look at how the phrase and its usage have evolved in popular culture, see the related piece titled what happened to fly like a bird. See the related piece 'fly like a bird uh huh what you heard' for more on how specific song lyrics and popular phrases shape cultural images of avian flight. There is even a line dance called 'Fly Like a Bird' with its own step-by-step choreography, a very different kind of 'flying bird step by step' that has nothing to do with pectoralis muscles but everything to do with the human desire to embody that freedom physically.
It is also worth addressing directly what 'shapeshifting into a bird' means in this context, since it appears as a related search. Physically transforming into a bird is not possible and belongs firmly in the realm of folklore, mythology, and fantasy literature, not biology. Human DNA does not contain the developmental programs for feathers, hollow bones, or a keeled sternum, and no known mechanism in biology allows somatic transformation of that kind. What shapeshifting stories represent, across cultures from Norse mythology to Indigenous American traditions, is a deep imaginative identification with birds and the qualities they symbolize: perspective, freedom, and the ability to transcend ordinary limits. That is a rich cultural conversation worth having, just not one to confuse with avian biomechanics.
Safety and ethical notes for bird watching and human flight
If this article has made you want to spend more time watching birds in flight, that impulse is worth acting on. Studying real birds in the field, with binoculars or a camera, is one of the best ways to internalize the flight phases described above. When observing nesting or roosting sites, maintain respectful distance (generally at least 30 meters for sensitive species) and avoid flushing birds repeatedly, as each forced takeoff carries genuine metabolic costs, especially during breeding season or cold weather. For human flight activities, always train with certified instructors, follow national aviation authority guidelines, and never rush your progression. The fatality statistics in paragliding and hang gliding drop dramatically for pilots who complete structured training and fly conservatively in their early years.
Putting it all together
Every time a bird launches from a branch, it is executing a precisely orchestrated sequence of muscular contractions, skeletal articulations, and aerodynamic events that took millions of years to refine. The six phases, from the explosive burst of takeoff through the delicate sensory ballet of landing, each involve different anatomical structures, different energy systems, and different aerodynamic regimes. Understanding that sequence step by step is not just an academic exercise. It gives you a new way of seeing every bird you watch, recognizing the alula deploying on a landing pigeon, noticing the asymmetric tail twist as a red-tailed hawk banks into a thermal, appreciating why a hummingbird is working so much harder than it looks. And if it inspires you to strap into a paraglider harness and find out firsthand what a thermal feels like rising beneath a wing, so much the better.
FAQ
What is the single core research question to answer when writing an article titled “flying bird step by step”?
How can I explain, accurately and accessibly, both (a) the biological step‑by‑step mechanics of bird flight (phases, anatomy, aerodynamics, energetics, and species differences) and (b) realistic, safe human ways to experience birdlike flight, while distinguishing scientific facts from cultural, metaphorical or fictional variants?
Which high‑level evidence sources are essential to define user/search intent for this query?
Combine: (1) Google Trends to identify seasonal/geo interest and common search variants (tier2); (2) Google Search Quality Rater Guidelines / SERP analysis to classify intent (Know/Do/Visit) and expected page types (tier1); and (3) commercial keyword tools (Ahrefs/SEMrush) to quantify volumes and label intent (commercial).
What primary biomechanics and modeling literature should I cite for phase definitions and energetic models?
Cite Colin J. Pennycuick’s Modelling the Flying Bird (2008) for canonical flight‑performance theory and power curves, plus classic Pennycuick migration/mechanics papers. Augment with review and synthesis papers in Journal of Experimental Biology that summarize phase sequencing and power/kinematic boundaries (tier1).
Which empirical kinematic and muscle physiology studies are necessary to link flight phases to wing motion and muscles?
Use high‑speed videography and 3‑D kinematics studies (e.g., Tobalske et al. for hummingbirds, J. Exp. Biol. kinematic papers) for stroke plane/wingbeat metrics, and EMG/muscle activation studies showing pectoralis (downstroke) and supracoracoideus (upstroke via pulley) timing and force production (tier1).
What fluid‑mechanics and aerodynamic sources explain how aerodynamic mechanisms change across flight phases and sizes?
Reference reviews in Annual Review of Fluid Mechanics on vortices and forces in biological flight and targeted J. Exp. Biol. papers on leading‑edge vortices (LEV), wake capture, induced drag and vortex wake topology. Include experimental CFD/particle image velocimetry (PIV) studies that show when quasi‑steady models break down (tier1).
How should I document phase‑specific energetics and provide quantitative estimates?
Use Pennycuick’s flight‑performance theory for power curves and complement with field metabolic studies (doubly labelled water, respirometry) and species field studies (e.g., albatross energetics) to show relative costs (takeoff/hover high cost; dynamic soaring very low per‑distance cost). Include citations to doubly labelled water field papers and Pennycuick classics (tier1).